Proteinojenik amino asit
Proteinojenik amino asitler, translasyon- proteinlerin sentezi- sürecine katılan amino asitlerdir. Proteinojenik kelimesi "protein oluşturan" manasına gelip "protein" ve "+jen(oluşturan, doğuran, ortaya çıkaran)" yapılarından türetilmiştir. Bilinen hayatta 22 adet genetik olarak kodlanmış(proteinojenik) amino asit mevcuttur. Bunlardan 20 tanesi genetik koda kodlanmış şekildeyken buna ek olarak 2 tanesi(Selenosistein ve Pirolizin)[a] özel translasyon mekanizmaları ile kullanılabilmektedir.[1]

Bunların zıttı olan non-proteinojenik amino asitler, proteinlerin yapısına katılmayan(mesela GABA, L-DOPA, Triiyodotironin), yanlışlıkla genetik olarak kodlanmış bir amino asidin yerine geçebilen veya standart hücre mekanizmasınca izolasyonda ve direkt üretilemeyen(mesela Hidroksiprolin) amino asitlere verilen isimdir. Bazı non-proteinojenik amino asitler non-ribozomik peptitlerin yapısına katılır.
Hem ökaryot hem de prokaryotlarda selenosistein protein yapısına katılabilmektedir. Selenosistein, SECIS elementi olarak bilinen bir nükleotid dizisi aracılığıyla yapıya katılabilmektedir. SECIS elementi normalde stop kodonu olan UGA dizinin[b] selenosistein şeklindesinde okunmasını sağlar. Bazı metanojenik prokaryotlarda normalde stop kodonu olan UAG kodonu[c] da pirolizin şeklinde okunabilir. Prolizinin proteinin yapısına katılma mekanizması tam olarak bilinmemektedir.[2]
Ökaryotlarda 20 genetik koda kodlanmış amino asit ve selenosistein olmak üzere toplam 21 tane amino asit kullanılmaktadır. İnsanlar bu amino asitlerin 12 tanesi birbirlerinden veya ara metabolizma ürinlerinden sentezleyebilir. Diğer 9 tanesini dışardan alması gerekir ve bunlara esansiyel amino asitler denir. Bu amino asitler; fenilalanin, histidin, izolösin, lizin, lösin, metiyonin, treonin, triptofan, valin idir.[3]
Proteinojenik amino asitlerin, ribozim otoaminoasilasyon sistemi tarafından tanınabilen amino asitlerle ilişki olduğu tespit edilmiştir.[4] Bundan, bu amino asitlerden olmayanların evrim sonucunda nükleotid-temelli yaşamdan dışlanıp non-proteinojenik amino asitler sınıfına katılması sebep olduğu iddia edilmiştir. Bazı spesifik non-proteinojenik amino asitlerin(mesela yarı ömrü nispeten kısa olan ornitin ve homoserin, toksik olan kanavanin) protein yapısına katılmamasına dair başka açıklamalar da sunulmuştur.[kaynak belirtilmeli]
Yapıları
değiştirAlttaki infografikte, ökaryotların genetik kodunda kodlanmış 21 amino asidin yapısı ve isimlerini kısaltması verilmiştir. Yapılar standart kimyasal yapıları olup sulu çözeltilerdeki hali olan zwitteriyon halinde değildir.

22 Amino asidin sköletik gösterimi(en) şöyledir:
-
 L-Alanin (Ala/A)
L-Alanin (Ala/A) -
 L-Arjinin (Arg/R)
L-Arjinin (Arg/R) -
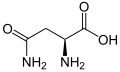 L-Asparajin (Asn/N)
L-Asparajin (Asn/N) -
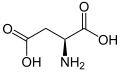 L-Aspartik asit (Asp/D)
L-Aspartik asit (Asp/D) -
 L-Sistein (Cys/C)
L-Sistein (Cys/C) -
 L-Glutamik asit (Glu/E)
L-Glutamik asit (Glu/E) -
 L-Glutamin (Gln/Q)
L-Glutamin (Gln/Q) -
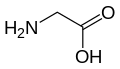 Glisin (Gly/G)
Glisin (Gly/G) -
 L-Histidin (His/H)
L-Histidin (His/H) -
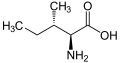 L-İzolösin (Ile/I)
L-İzolösin (Ile/I) -
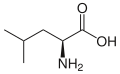 L-Lösin (Leu/L)
L-Lösin (Leu/L) -
 L-Lizin (Lys/K)
L-Lizin (Lys/K) -
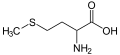 L-Metiyonin (Met/M)
L-Metiyonin (Met/M) -
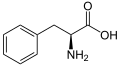 L-Fenilalanin (Phe/F)
L-Fenilalanin (Phe/F) -
 L-Prolin (Pro/P)
L-Prolin (Pro/P) -
 L-Serin (Ser/S)
L-Serin (Ser/S) -
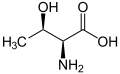 L-Treonin (Thr/T)
L-Treonin (Thr/T) -
 L-Triptofan (Trp/W)
L-Triptofan (Trp/W) -
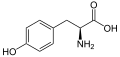 L-Tirozin (Tyr/Y)
L-Tirozin (Tyr/Y) -
 L-Valin (Val/V)
L-Valin (Val/V)


















IUPAC/IUBMB, aşağıdaki iki amino asit için kullanılan kısaltmaları önermektedir:
-
 L-Selenosistein (Sec/U)
L-Selenosistein (Sec/U) -
 L-Pirolizin (Pyl/O)
L-Pirolizin (Pyl/O)

Kimyevi özellikleri
değiştirAşağıdaki tablolarda amino asitlerin kısaltmalarını, amino asitlerin ve yan zincirlerinin kimyevi hususiyetlerine dair bilgiler listelenmiştir. Doğadaki bolluklarına nispetle izotoplarının ortalama ağırlığı hesaplanmıştır. Oluşan her peptit bağı sonucunda molekül bir su molekülü kaybeder. Bundan dolayı hesaplalarda her peptit bağı için 18,01524 Da molekül ağırlığından çıkarılır.
Kimyasal özellikler
değiştir| Amino asit | Kısaltmaları | Ortalama ağırlık (Da) | pI | pK1
(α-COO-) |
pK2
(α-NH3+) |
|---|---|---|---|---|---|
| Alanin | A/Ala | 89,09404 | 6,01 | 2,35 | 9,87 |
| Sistein | C/Cys | 121,15404 | 5,05 | 1,92 | 10,70 |
| Aspartik asit | D/Asp | 133,10384 | 2,85 | 1,99 | 9,90 |
| Glutamik asit | E/Glu | 147,13074 | 3,15 | 2,10 | 9,47 |
| Fenilalanin | F/Phe | 165,19184 | 5,49 | 2,20 | 9,31 |
| Glisin | G/Gly | 75,06714 | 6,06 | 2,35 | 9,78 |
| Histidin | H/His | 155,15634 | 7,60 | 1,80 | 9,33 |
| İzolösin | I/Ile | 131,17464 | 6,05 | 2,32 | 9,76 |
| Lizin | K/Lys | 146,18934 | 9,60 | 2,16 | 9,06 |
| Lösin | L/Leu | 131,17464 | 6,01 | 2,33 | 9,74 |
| Metiyonin | M/Met | 149,20784 | 5,74 | 2,13 | 9,28 |
| Asparajin | N/Asn | 132,11904 | 5,41 | 2,14 | 8,72 |
| Pirolizin | O/Pyl | 255,31 | ? | ? | ? |
| Prolin | P/Pro | 115,13194 | 6,30 | 1,95 | 10,64 |
| Glutamin | Q/Gln | 146,14594 | 5,65 | 2,17 | 9,13 |
| Arjinin | R/Arg | 174,20274 | 10,76 | 1,82 | 8,99 |
| Serin | S/Ser | 105,09344 | 5,68 | 2,19 | 9,21 |
| Treonin | T/Thr | 119,12034 | 5,60 | 2,09 | 9,10 |
| Selenosistein | U/Sec | 168,053 | 5,47 | 1,91 | 10 |
| Valin | V/Val | 117,14784 | 6,00 | 2,39 | 9,74 |
| Triptofan | W/Trp | 204,22844 | 5,89 | 2,46 | 9,41 |
| Tirozin | Y/Tyr | 181,19124 | 5,64 | 2,20 | 9,21 |
Yan dal özellikleri
değiştir| Amino asit | Tek harfi kısatlması | Üç harfli kısatlması | Yan dal | Hidrofobik | pKa[d] | Polar | pH | Küçük | Ufak | Aromatikveya Alifatik | Van der Waals hacmi (Å3) |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Alanin | A | Ala | -CH3 | - | - | alifatik | 67 | ||||
| Sistein | C | Cys | -CH2SH | 8,55 | asidik | - | 86 | ||||
| Aspartik asit | D | Asp | -CH2COOH | 3,67 | asidik | - | 91 | ||||
| Glutamik asit | E | Glu | -CH2CH2COOH | 4,25 | asidik | - | 109 | ||||
| Fenilalanin | F | Phe | -CH2C6H5 | - | - | aromatik | 135 | ||||
| Glisin | G | Gly | -H | - | - | - | 48 | ||||
| Histidin | H | His | -CH2-C3H3N2 | 6,54 | zayıf bazik | aromatik | 118 | ||||
| İzolösin | I | Ile | -CH(CH3)CH2CH3 | - | - | alifatik | 124 | ||||
| Lizin | K | Lys | -(CH2)4NH2 | 10,40 | bazik | - | 135 | ||||
| Lösin | L | Leu | -CH2CH(CH3)2 | - | - | alifatik | 124 | ||||
| Metiyonin | M | Met | -CH2CH2SCH3 | - | - | alifatik | 124 | ||||
| Asparajin | N | Asn | -CH2CONH2 | - | - | - | 96 | ||||
| Pirolizin | O | Pyl | -(CH2)4NHCOC4H5NCH3 | Veri yok | zayıf bazik | - | ? | ||||
| Prolin | P | Pro | -CH2CH2CH2- | - | - | - | 90 | ||||
| Glutamin | Q | Gln | -CH2CH2CONH2 | - | - | - | 114 | ||||
| Arjinin | R | Arg | -(CH2)3NH-C(NH)NH2 | 12,3 | güçlü bazik | - | 148 | ||||
| Serin | S | Ser | -CH2OH | - | - | - | 73 | ||||
| Treonin | T | Thr | -CH(OH)CH3 | - | - | - | 93 | ||||
| Selenosistein | U | Sec | -CH2SeH | 5,43 | asidik | - | ? | ||||
| Valin | V | Val | -CH(CH3)2 | - | - | alifatik | 105 | ||||
| Triptofan | W | Trp | -CH2C8H6N | - | - | aromatik | 163 | ||||
| Tirozin | Y | Tyr | -CH2-C6H4OH | 9,84 | zayıf asidik | aromatik | 141 |
Not: Amino asit residülerinin küçük peptidlerdeki pKa değerleri, proteinlerin içinde bir miktar farklılık gösterir. Protein pKa hesaplaması(en) bu aradaki farkı hesaplamak için kullanılabilir,
Gen ekspresyonu ve biyokimya
değiştir| Amino asit | Tek harfli kısaltması | Üç harfli kısaltması | Kodon(lar) | Mevcudiyet | İnsan için esansiyelliğie | |||
|---|---|---|---|---|---|---|---|---|
| Arke proteinlerinde (%)f |
Bakteri proteinlerinde (%)f |
Ökaryot proteinlerinde (%)f |
İnsan proteinlerinde (%)f | |||||
| Alanin | A | Ala | GCU, GCC, GCA, GCG | 8,2 | 10,06 | 7,63 | 7,01 | Hayır |
| Sistein | C | Cys | UGU, UGC | 0,98 | 0,94 | 1,76 | 2,3 | Kondisyonel |
| Aspartik asit | D | Asp | GAU, GAC | 6,21 | 5,59 | 5,4 | 4,73 | Hayır |
| Glutamik asit | E | Glu | GAA, GAG | 7,69 | 6,15 | 6,42 | 7,09 | Kondisyonel |
| Fenilalanin | F | Phe | UUU, UUC | 3,86 | 3,89 | 3,87 | 3,65 | Evet |
| Glisin | G | Gly | GGU, GGC, GGA, GGG | 7,58 | 7,76 | 6,33 | 6,58 | Kondisyonel |
| Histidin | H | His | CAU, CAC | 1,77 | 2,06 | 2,44 | 2,63 | Evet |
| İzolösin | I | Ile | AUU, AUC, AUA | 7,03 | 5,89 | 5,1 | 4,33 | Evet |
| Lizin | K | Lys | AAA, AAG | 5,27 | 4,68 | 5,64 | 5,72 | Evet |
| Lösin | L | Leu | UUA, UUG, CUU, CUC, CUA, CUG | 9,31 | 10,09 | 9,29 | 9,97 | Evet |
| Metiyonin | M | Met | AUG | 2,35 | 2,38 | 2,25 | 2,13 | Evet |
| Asparajin | N | Asn | AAU, AAC | 3,68 | 3,58 | 4,28 | 3,58 | Hayır |
| Pirolizin | O | Pyl | UAGa | 0 | 0 | 0 | 0 | Hayır |
| Prolin | P | Pro | CCU, CCC, CCA, CCG | 4,26 | 4,61 | 5,41 | 6,31 | Hayır |
| Glutamin | Q | Gln | CAA, CAG | 2,38 | 3,58 | 4,21 | 4,77 | Hayır |
| Arjinin | R | Arg | CGU, CGC, CGA, CGG, AGA, AGG | 5,51 | 5,88 | 5,71 | 5,64 | Kondisyonel |
| Serin | S | Ser | UCU, UCC, UCA, UCG, AGU, AGC | 6,17 | 5,85 | 8,34 | 8,33 | Hayır |
| Treonin | T | Thr | ACU, ACC, ACA, ACG | 5,44 | 5,52 | 5,56 | 5,36 | Evet |
| Selenosistein | U | Sec | UGAb | 0 | 0 | 0 | >0 | Hayır |
| Valin | V | Val | GUU, GUC, GUA, GUG | 7,8 | 7,27 | 6,2 | 5,96 | Evet |
| Triptofan | W | Trp | UGG | 1,03 | 1,27 | 1,24 | 1,22 | Evet |
| Tirozin | Y | Tyr | UAU, UAC | 3,35 | 2,94 | 2,87 | 2,66 | Kondisyonel |
| Stop kodonuc | - | Term | UAA, UAG, UGAd | ? | ? | ? | — | — |
a UAG normalde amber stop kodonu olsa da pylTSBCD gen kümesine kodlanmış biyolojik mekanizmaya sahip olan hücrelerde pirolizinin yapıya katılmasını sağlar.[8]
b UGA normale opal stop kodonu olsa da SECIS elementi olarak bilinen bir nükleotid dizisi aracılığıyla selenosisteinin yapıya katılmasını temin etmektedir.
c Stop kodonu herhangi bir amino asit kodlamasa da bütünlük için tabloya eklenmiştir.
d UAG ve UGA her zaman stop kodonu değildir (yukarıya bkz.).
e Esansiyel amino asitler insanda sentezlenemeyen ve dışarıdan alınması gereken amino asitlerdir. Kondisyonel(şartlı) amino asitler normalde dışarıdan alınması gerekmese de yeterli miktarda sentez yapamayan spesifik popülasyonun egzojenik(en) takviyeye ihtiyacı vardır.
f Amino asitlerin mevcudiyet miktarı 135 arke, 3776 bakteri, 614 ökaryot proteomu[9] ve insan proteomuna(21.006 protein)[kaynak belirtilmeli] dayanmaktadır.
Kütle spektrometrisi
değiştirKütle spektrometrisinin peptit ve proteinlerde kulanımı ile residülerin[e] kütleleri hesaplanabilmektedir. Protein veya peptitlerin toplam kütlesi, residülerin ve suyun (Monoizotopik kütle = 18,01056 Da; Ortalama kütle = 18,0153 Da) kütlesinin toplamından ibarettir. Aşağıda tablolandırılmış residülerin kütlesi kimyasal formül ve atomik ağırlıkları ile hesaplanmıştır.[10] Kütle spektrometrisinde iyonlar ayrıca bir veya birden fazla proton içerebilir (Monoizotopik kütle = 1,00728 Da; Ortalama kütle[f] = 1,0074 Da).
| Amino asit | Tek harfli kısaltması | Üç harfli kısaltması | Formül | Mon. mass
(Da) |
Avg. mass (Da) |
|---|---|---|---|---|---|
| Alanin | A | Ala | C3H5NO | 71,03711 | 71,0779 |
| Sistein | C | Cys | C3H5NOS | 103,00919 | 103,1429 |
| Aspartik asit | D | Asp | C4H5NO3 | 115,02694 | 115,0874 |
| Glutamik asit | E | Glu | C5H7NO3 | 129,04259 | 129,1140 |
| Fenilalanin | F | Phe | C9H9NO | 147,06841 | 147,1739 |
| Glisin | G | Gly | C2H3NO | 57,02146 | 57,0513 |
| Histidin | H | His | C6H7N3O | 137,05891 | 137,1393 |
| İzolösin | I | Ile | C6H11NO | 113,08406 | 113,1576 |
| Lizin | K | Lys | C6H12N2O | 128,09496 | 128,1723 |
| Lösin | L | Leu | C6H11NO | 113,08406 | 113,1576 |
| Metiyonin | M | Met | C5H9NOS | 131,04049 | 131,1961 |
| Asparajin | N | Asn | C4H6N2O2 | 114,04293 | 114,1026 |
| Pirolizin | O | Pyl | C12H19N3O2 | 237,14773 | 237,2982 |
| Prolin | P | Pro | C5H7NO | 97,05276 | 97,1152 |
| Glutamin | Q | Gln | C5H8N2O2 | 128,05858 | 128,1292 |
| Arjinin | R | Arg | C6H12N4O | 156,10111 | 156,1857 |
| Serin | S | Ser | C3H5NO2 | 87,03203 | 87,0773 |
| Treonin | T | Thr | C4H7NO2 | 101,04768 | 101,1039 |
| Selenosistein | U | Sec | C3H5NOSe | 150,95364 | 150,0489 |
| Valin | V | Val | C5H9NO | 99,06841 | 99,1311 |
| Triptofan | W | Trp | C11H10N2O | 186,07931 | 186,2099 |
| Tirosin | Y | Tyr | C9H9NO2 | 163,06333 | 163,1733 |
Stokiyometri ve hücrenin metabolik maliyeti
değiştirAşağıda E.coli hücresindeki amino asitlerin bolluğu ve bu amino asitlerin sentezindeki metabolik maliyeti(ATP) tablolaştırılmıştır. Sentezdeki ATP maliyeti kısmındaki negatif sayılar metabolik süreçte net enerji açığa çıktığını ve hücreye ATP maliyetinin olmadığını bildirir.[11] Amino asitlerin bolluk miktarına serbest halleri ve polimerize halleri(proteinler) dahildir.[kaynak belirtilmeli]
| Amino asit | Tek harfli kısaltması | Üç harfli kısaltması | Bolluk (E. coli hücresi başına # molekülünün sayısı(×108) ) |
Sentezindeki ATP maliyeti | |
|---|---|---|---|---|---|
| Aerobik kondüsyon |
Anaerobik kondüsyon | ||||
| Alanin | A | Ala | 2,9 | -1 | 1 |
| Sistein | C | Cys | 0,52 | 11 | 15 |
| Aspartik asit | D | Asp | 1,4 | 0 | 2 |
| Glutamik asit | E | Glu | 1,5 | -7 | -1 |
| Fenilalanin | F | Phe | 1,1 | -6 | 2 |
| Glisin | G | Gly | 3,5 | -2 | 2 |
| Histidin | H | His | 0,54 | 1 | 7 |
| İzolösin | I | Ile | 1,7 | 7 | 11 |
| Lizin | K | Lys | 2,0 | 5 | 9 |
| Lösin | L | Leu | 2,6 | -9 | 1 |
| Metiyonin | M | Met | 0,88 | 21 | 23 |
| Asparajin | N | Asn | 1,4 | 3 | 5 |
| Pirolizin | O | Pyl | - | - | - |
| Prolin | P | Pro | 1,3 | -2 | 4 |
| Glutamin | Q | Gln | 1,5 | -6 | 0 |
| Arjinin | R | Arg | 1,7 | 5 | 13 |
| Serin | S | Ser | 1,2 | -2 | 2 |
| Treonin | T | Thr | 1,5 | 6 | 8 |
| Selenosistein | U | Sec | - | - | - |
| Valin | V | Val | 2,4 | -2 | 2 |
| Triptofan | W | Trp | 0,33 | -7 | 7 |
| Tirozin | Y | Tyr | 0,79 | -8 | 2 |
Şerh
değiştir| Amino asit | Kısaltması | Açıklama | |
|---|---|---|---|
| Alanin | A | Ala | Oldukça bol ve versatildir. Glisinden daha serttir. Protein konformasyonunda ufak sterik limitleri(en) oluşturabilecek küçüklüktedir. Hemen hemen nötral karakterdedir ve hem proteinin dışındaki hidrofilik bölgede hem de proteinin iç kısmındaki hidrofobik kısımda bulunabilir. |
| Asparajin veya aspartik asit | B | Asx | Bu iki amino asidin de olabileceği yerlerin adıdır. |
| Sistein | C | Cys | Ağır metallere kolayca bağlanabilen bir sülfür atomuna sahiptir. Okside kondisyondayken iki sistein disülfit bağıyla birleşerek sistin adlı dimerik amino asidi oluşturur. Sistin bir proteine katıldığında, mesela insülin, üçüncül yapıyı stabilize ederek denatürasyona daha dayanıklı yapar. Bu yüzden disülfat bağları sert ortamlarda faaliyette olan proteinlede (mesela pepsin, kimotripsin gibi sindirim enzimlerinde ve keratin gibi strüktürel proteinlerde) yaygındır. Disülfit bağları insülin gibi tek başına stabil olmayacak kadar küçük peptitlerde de bulunur. |
| Aspartik asit | D | Asp | Glutamik aside benzer bir davranışı vardır ve güçlü negatif yüklü hidrofilik ve asidik bir yan zincire sahiptir. Genellikle proteinlerin dış kısmında olarak proteini suda çözünür kılar. Pozitif yüklü molekül ve iyonlara bağlanır. Sıklıkla enzimlerdeki metal iyonlarını sabitleyici bir rolü vardır. İç kısımda bulunduğu çoğu halde arjinin ve lizin gibi bir pozitif yüklü amino asitle eşleşmiştir. |
| Glutamik asit | E | Glu | Aspartik aside benzer bir davranışı vardır ve aspartik aside kıyasla daha uzun ve esnek bir yan zincire sahiptir. |
| Fenilalanin | F | Phe | Hayvanlar için esansiyeldir. Fenilalanin, tirozin ve triptofan büyük, rijit aromatik yan dallara sahiptir. Bunlar en büyük amino asitlerdir. İzolösin, lösin ve valin gibi bu amino asitler de hidrofobiik olduklarından katlanmış proteinin iç kısmında yer almaya meyilli olurlar. Fenilalanin tirozine dönüşebilir. |
| Glisin | G | Gly | İki hidrojen atomu α karbona bağlı olduğundan şiral değil yani enantiyomeri yoktur. En küçük amino asittir. Protein zincirinde kolayca rotatif hareket yapabilmekte ve zincire esneklik kazandırmaktadır. Oldukça sıkı alanlara dahi sığabilmektedir. Mesela kollajenin üçlü sarmal yapısı. Lakin strüktürel gibi yapılarda çok fazla esneklik genellikle arzulanmadığından alaninden daha azdır. |
| Histidin | H | His | Hayvanlar için esansiyeldir. Hafif asidik bir ortamda dahi nitrojenler protonasyona uğrar ki bu da histidinin ve tüm polipeptidin özelliklerini değiştirir. Çoğu düzenleyici mekanizmalarda kullanılmaktadır. Mesela geç endozom veya lizozomdaki enzimlerin düzenlenmesini sağlar çünkü polipeptidin konformasyonu ve davranışı asidik ortamlarda değiştirmektedir. Lakin bunun için gereken histidin sayısı azdır. Bundan dolayı histidin miktarı nispeten oldukça azdır. |
| İzolösin | I | Ile | İzolösin hayvanlar için esansiyeldir. İzolösin, lösin ve valine büyük bir alifatik hidrofobik yan zincire sahiptir. Rijit molekülerdir ve karşılıklı hidrofobik etkileşimleri proteinlerin doğru katlanması için oldukça önemlidir çünkü bu amino asitlerin bulunduğu zincir kısımları proteinin iç kısmında yer almaya meyilli olur. |
| Lösin vya izolösin | J | Xle | Bu iki amino asidin de olabileceği yerlerin adıdır. |
| Lizin | K | Lys | Hayvanlar için esansiyeldir. Arjinine benzer bir karakteristiktedir. Ucu pozitif yüklü, uzun ve esnek bir yan zincire sahiptir. Yan zincirlerinin esnekliği lizin ve arjinini çoğu negatif yüklü molekülün yüzeyine bağlanmasını sağlar. Mesela DNA-bağlayan proteinlerin(örneğin histon) aktif bölgesi bu yüzden arjinin ve lizin bakımından zengindir. Sahip oldukları güçlü pozitif yük, bu amino asitleri dış hidrofilik yüzeyde bulunmaya meyilli yapar. İç kısımda bulundukları çoğu zamandaysa aspartat ve glutamat gibi bir negatif yüklü amino asitle eşleşmişlerdir. |
| Lösin | L | Leu | Lösin hayvanlar için esansiyeldir. İzolösin ve valine benzer bir karakterdedir. |
| Metiyonin | M | Met | Hayvanlar için esansiyeldir. Her zaman proteinin ilk amino asididir ve bazan translasyon sonrasında proteinden çıkarılır. Sistein gibi bir sülfüre sahiptir lakin sülfüre hidrojen yerine metil grubu merbuttur. Metil grubu stimüle edilerek çoğu reaksiyonda başka moleküllere yeni karbon atomu olarak katılmaktadır. |
| Asparajin | N | Asn | Aspartik aside benzerdir. Asparajin, aspartik asitte karboksil grubu bulunan yerde amid grubuna sahiptir. |
| Pirolizin | O | Pyl | Lizine benzerdir ama bir pirolin halkasına sahiptir. |
| Prolin | P | Pro | Prolin, N-terminaline bağlanmış istisnai bir halka yapısına sahiptir. Bu özelliği onu sabit bir CO-NH amid dizisi konformasyonda tutar. α-heliks veya β yaprak gibi katlanan proteinlerin yapısını bozabilmekte ve protein zincirini kink oluşturmaya zorlamaktadır. Kollajenlerde yaygındr ve sıklıkla posttranslasyonel modifikasyon sonucuda hidroksiproline dönüşür. |
| Glutamin | Q | Gln | Glutamik aside benzer olup glutamik asitte karboksil olduğu yerde amid grubuna sahiptir. Proteinlerin yapımında ve amonyağın depolanmasında kullanılır ve vücutta en bol bulunan amino asittir. |
| Arjinin | R | Arg | Lizine benzer bir fonksiyonu vardır. |
| Serin | S | Ser | Serin ve treonin ucunda hidroksil grubu kısa bir yan dala sahiptir. Bu hidroksil grubu hidrojen vermeye meyilli olduğundan dolayı serin ve treonin enzimlerde genellikle hidrojen donörü olarak davranır. İkisi de oldukça hidrofiliktir ve bundan dolayı suda çözünebilir proteinlerin dış kısımları bu amino asitler bakımından zengin olmaya meyillidir. |
| Treonin | T | Thr | Treonin insanlar için elzem(esansiyel) bir amino asittir. Serine benzer bir karakterdedir. |
| Selenosistein | U | Sec | Sisteinin kükürt yerine selenyumlu halidir. |
| Valin | V | Val | Lösin hayvanlar için esansiyeldir. İzolösin ve lösine benzer bir karakterdedir. |
| Triptofan | W | Trp | Triptofan hayvanlar için esansiyeldir. Fenilalanin ve tirozine benzer bir karakteristiği vardır. Serotoninin prekürsörü ve doğal floresanstır. |
| Bilinmeyen/Meçhul | X | Xaa | Amino asidin bilinmediği önemsiz yerlerin adıdır. |
| Tirozin | Y | Tyr | Tirozin, fenilalanin (tirozinin prekürsörü) ve triptofana benzer karakteristiktedir. Tirozin melanin, epinefrin ve tiroit hormonlarının prekürsörüdür. Doğal floresanstır ve floresansı genellikle triptofanlara enerjisinin transferi ile diner. |
| Glutamik asit veya glutamin | Z | Glx | Bu iki amino asidin de olabileceği yerlerin adıdır. |
Katabolizma
değiştir
Amino asitler ana ürünlerinin özelliklerine göre tasnif edilebilirler:[12]
- Glukojenikler, glukoneogenez ile glükoz hasıl etme yeteneğine sahip ürünlere dönüşenlerdir.
- Ketojenikler, ürünlerinin glükoz oluşturma yeteneği olmayanlardır. Ürünleri ketogenez veya lipit sentezinde kullanılabilir.
- Hem glukojenik hem de ketojenik ürünlere katalizlenebilenlerdir.
Ayrıca bakınız
değiştirDipnot
değiştir- ^ Bunlara non-kanonikal amino asitler denmektedir.
- ^ Nam-ı diğer opal kodonu
- ^ Nam-ı diğer amber kodonu
- ^ Asp, Cys, Glu, His, Lys ve Tyr'nin değerleri alanin pentapeptidin merkezine yerleştirilmiş amino asitler kullanılarak belirlenmiştir.[5] Arg'nin değeri Paceet et al. (2009)'den alınmıştır.[6] Sec'in değeri Byun & Kang (2011)'den alınmıştır.[7]
- ^ Residü yani kalıntıdan kasıt amino asittir.
- ^ Protonların ortalama bir kütlesi olamaz. Bu kafa karıştırıcı istidlal, döteronun cari bir izotop olduğunu gösterse de bunlar farklı bir tür olmalıdır(bkz. Hidron (kimya)).
Kaynakça
değiştir- ^ Ambrogelly, Alexandre; Palioura, Sotiria; Söll, Dieter (Ocak 2007). "Natural expansion of the genetic code". Nature Chemical Biology (İngilizce). 3 (1): 29-35. doi:10.1038/nchembio847. ISSN 1552-4469. 16 Ocak 2025 tarihinde kaynağından arşivlendi. Erişim tarihi: 6 Ocak 2025.
- ^ Lobanov, Alexey V.; Turanov, Anton A.; Hatfield, Dolph L.; Gladyshev, Vadim N. (1 Ağustos 2010). "Dual functions of codons in the genetic code". doi:10.3109/10409231003786094?scroll=top&needaccess=true. ISSN 1040-9238.
- ^ Young, Vernon R (1 Ağustos 1994). "Adult Amino Acid Requirements: The Case for a Major Revision in Current Recommendations1". The Journal of Nutrition. 124: 1517S-1523S. doi:10.1093/jn/124.suppl_8.1517S. ISSN 0022-3166.
- ^ Erives, Albert (1 Ağustos 2011). "A Model of Proto-Anti-Codon RNA Enzymes Requiring l-Amino Acid Homochirality". Journal of Molecular Evolution (İngilizce). 73 (1): 10-22. doi:10.1007/s00239-011-9453-4. ISSN 1432-1432. PMC 3223571 $2. PMID 21779963. 20 Eylül 2018 tarihinde kaynağından arşivlendi. Erişim tarihi: 10 Ocak 2025.
- ^ Thurlkill, Richard L.; Grimsley, Gerald R.; Scholtz, J. Martin; Pace, C. Nick (2006). "pK values of the ionizable groups of proteins". Protein Science (İngilizce). 15 (5): 1214-1218. doi:10.1110/ps.051840806. ISSN 1469-896X. PMC 2242523 $2. PMID 16597822. 8 Ocak 2025 tarihinde kaynağından arşivlendi. Erişim tarihi: 11 Ocak 2025.
- ^ Pace, C. Nick; Grimsley, Gerald R.; Scholtz, J. Martin (15 Mayıs 2009). "Protein ionizable groups: pK values and their contribution to protein stability and solubility". The Journal of Biological Chemistry. 284 (20): 13285-13289. doi:10.1074/jbc.R800080200. ISSN 0021-9258. PMC 2679426 $2. PMID 19164280. 23 Nisan 2024 tarihinde kaynağından arşivlendi. Erişim tarihi: 11 Ocak 2025.
- ^ Byun, Byung Jin; Kang, Young Kee (Mayıs 2011). "Conformational preferences and p Ka value of selenocysteine residue". Biopolymers (İngilizce). 95 (5): 345-353. doi:10.1002/bip.21581. ISSN 0006-3525. 30 Kasım 2022 tarihinde kaynağından arşivlendi. Erişim tarihi: 11 Ocak 2025.
- ^ Rother, Michael; Krzycki, Joseph A. (17 Ağustos 2010). "Selenocysteine, pyrrolysine, and the unique energy metabolism of methanogenic archaea". Archaea (Vancouver, B.C.). 2010: 453642. doi:10.1155/2010/453642. ISSN 1472-3654. PMC 2933860 $2. PMID 20847933. 30 Kasım 2024 tarihinde kaynağından arşivlendi. Erişim tarihi: 11 Ocak 2025.
- ^ Kozlowski, Lukasz P. (4 Ocak 2017). "Proteome-pI: proteome isoelectric point database". Nucleic Acids Research. 45 (D1): D1112-D1116. doi:10.1093/nar/gkw978. ISSN 1362-4962. PMC 5210655 $2. PMID 27789699. 19 Kasım 2024 tarihinde kaynağından arşivlendi. Erişim tarihi: 11 Ocak 2025.
- ^ "Atomic Weights and Isotopic Compositions for All Elements". physics.nist.gov. 18 Aralık 2024 tarihinde kaynağından arşivlendi. Erişim tarihi: 10 Ocak 2025.
- ^ Phillips R, Kondev J, Theriot J, Garcia HG, Orme N (2013). Physical biology of the cell (İkinci bas.). Garland Science. s. 178. ISBN 978-0-8153-4450-6.
- ^ Ferrier DR (2005). "Chapter 20: Amino Acid Degradation and Synthesis". Champe PC, Harvey RA, Ferrier DR (Ed.). Lippincott's Illustrated Reviews: Biochemistry (Lippincott's Illustrated Reviews) (İngilizce). Hagerstwon, MD: Lippincott Williams & Wilkins. ISBN 978-0-7817-2265-0.
Genel kaynakça
değiştir- Nelson, David L.; Cox, Michael M. (2000). Lehninger Principles of Biochemistry (3. edisyon). Worth Publishers. ISBN 978-1-57259-153-0.
- Kyte J, Doolittle RF (Mayıs 1982). "A simple method for displaying the hydropathic character of a protein". Journal of Molecular Biology. 157 (1): 105-32. CiteSeerX 10.1.1.458.454. doi:10.1016/0022-2836(82)90515-0. PMID 7108955.
- Meierhenrich, Uwe J. (2008). Amino acids and the asymmetry of life (1. edisyon). Springer. ISBN 978-3-540-76885-2.
- Biochemistry, Harpers (2015). Harpers Illustrated Biochemistry (30st ed.). Lange. ISBN 978-0-07-182534-4.